|
Coautores
Adriana María Salazar Montes* Juan Armendáriz**
Biologo, Universidad de Guadalajara, Guadalajara, México*
Bioquimico, Instituto de Biología Molecular en Medicina y Terapia Génica. Universidad de Guadalajara, Guadalajara, México**
|
|
Recepción del artículo: 0 de , 0000
|
|
Aprobación: 13 de marzo, 2015
|
 Conclusión breve
Conclusión breve
El envío de genes terapéuticos a células blanco en terapia génica requiere de un vehículo llamado vector, de los cuales los adenovirus son los más utilizados en protocolos clínicos.
Resumen
El envío de genes terapéuticos a células blanco en terapia génica requiere de un vehículo llamado vector, de los cuales los adenovirus (AD) son los más utilizados en protocolos clínicos. Si bien los AD son virus ADN poco patógenos para el hombre, pueden infectar los tractos respiratorio y gastrointestinal sin mayores complicaciones. Presentan varias características que los hacen los vectores ideales para su utilización en terapia génica. Entre las características con las que cuentan, destacan su amplio tropismo celular, su capacidad infectiva en cualquier etapa del ciclo celular, el tamaño del transgen -que puede ser de hasta 35 kb- y la relativa facilidad de producción y titulación en el laboratorio. Sin embargo, tienen como desventaja que la administración de AD a un organismo induce una respuesta inmune que impacta en la eficiencia de transducción, el tiempo de expresión del transgen y la eliminación de las células transducidas. En el presente artículo se describen las características de los AD como vectores virales y la respuesta inmune desarrollada en el organismo; asimismo, se analizan los posibles mecanismos moleculares por los cuales los AD despiertan una respuesta inmune innata como una respuesta adaptativa ante su presencia, así como las diversas estrategias utilizadas por diferentes autores para evadir esa respuesta inmune y evitar su pronta eliminación en las células transducidas.
Palabras clave
terapia génica, vectores virales, adenovirus, respuesta inmune innata,, respuesta inmune adaptativa.
Clasificación en siicsalud
Artículos originales> Expertos del Mundo>
página www.siicsalud.com/des/expertos.php/142470
Especialidades
 Principal: Genética Humana, Inmunología,
Principal: Genética Humana, Inmunología,
Relacionadas: Bioquímica, Farmacología, Infectología,
Enviar correspondencia a:
Adriana Salazar Montes, 44340, Guadalajara, México
The immune response is the worst enemy of adenovirus-based gene therapy
Abstract
Sending therapeutic genes to target cells in gene therapy requires a vehicle called vector, of which adenoviruses (Ad) are most commonly used in clinical protocols. Ads are DNA viruses that are not pathogenic for humans but which infect both respiratory and gastrointestinal tracts without complications. Ads have several features that make them ideal vectors for use in gene therapy. Those characteristics include broad cell tropism, infectivity at any cell cycle stage, size of transgene of up to 35 kb, and easy production and titration in the laboratory. Nevertheless, they present the enormous disadvantage that Ad administration to an organism induces a high immune response that impacts transduction efficiency, transgene expression time and the elimination of transduced cells. This article describes the characteristics of adenoviral vectors, the immune response to them and the possible molecular mechanisms through which adenoviruses awaken an innate and adaptive immune response, as well as the various strategies used by different authors to avoid the immune response and prevent early elimination of the transduced cells.
Key words
gene therapy, viral vectors, adenovirus, innate immune response, adaptive immune response
EN LA TERAPIA GÉNICA CON ADENOVIRUS LA RESPUESTA INMUNE ES EL PRINCIPAL ENEMIGO
|
(especial para SIIC © Derechos reservados) |
|
Artículo completo
Introducción
La terapia génica es una relativamente nueva alternativa de la biomedicina para el tratamiento de enfermedades. Consiste en el envío de material genético exógeno a células diana, con el objeto de retrasar la progresión de una enfermedad o corregir un defecto específico. Las enfermedades abordadas con terapia génica deben cumplir con ciertos criterios designados por comités médicos internacionales, con el fin de que se autorice el empleo de este tipo de terapia en ensayos clínicos. Entre los criterios más importantes se encuentran los siguientes: no debe existir un tratamiento efectivo para su curación; debe contar con un sustento científico sólido que justifique el tratamiento; debe existir información científica que avale que su uso se verá reflejado en una mejora para el paciente.1
Para la transferencia de genes a un órgano blanco se utiliza un vehículo llamado vector, de los cuales existen virales y no virales. Los vectores no virales son vehículos como liposomas, plásmidos, dextrán y fosfato de calcio. Los vectores virales, por su parte, son virus modificados a los que se les han eliminado sus genes patógenos y los necesarios para la replicación y, en su lugar, se coloca el gen terapéutico. Entre éstos se encuentran los retrovirus, los Ad, los virus adeno-asociados y los herpes virus, de los cuales los Ad son los más utilizados debido a que presentan ciertas ventajas sobre los otros vectores: amplio tropismo celular, capacidad infectiva en cualquier etapa del ciclo celular, tamaño del transgen de hasta 35 kilobases (kb) y relativa facilidad de producción y titulación en el laboratorio. Un Ad convertido en vector es llamado Ad recombinante (rAd).2 Hasta 2014 se tiene el registro de 1 996 ensayos clínicos a nivel mundial, de los cuales el porcentaje más alto (23.4%) utilizó un rAd para el envío de sus genes terapéuticos.3
Adenovirus
El genoma de los Ad es ADN de doble cadena de 26 a 44 kb. Existen 51 serotipos en los seres humanos, de los cuales Ad2 y Ad5 son los más utilizados por ser los mejor caracterizados, sin representar ningún riesgo para la salud. Los Ad contienen una cápside externa, un núcleo proteico central que recubre al ADN lineal y varias proteínas accesorias. La cápside tiene una conformación icosahédrica, donde 240 de sus capsómeros son hexones y 12 son pentones.4 Uno de estos pentones constituye la proteína fibrilar donde se encuentra la fracción globular knob, con la cual el Ad interactúa con el receptor CAR (receptor coxanquie-adenovirus) de la célula huésped para introducirse en ella.5
Los Ad empleados en terapia génica pueden ser de primera, segunda y tercera generación (estos últimos llamados también gutless).6 Los Ad de primera generación presentan deleciones en las regiones E1 y E3 de su genoma (necesarias para la replicación) y en su lugar se inserta el gen terapéutico. Los de segunda generación tienen suprimidas las regiones E1 a E4, mientras que en los de tercera generación o gutless la mayor parte del genoma es suprimido, por lo que pueden albergar genes más grandes y resultan menos inmunogénicos.6
A pesar de las ventajas mencionadas, los Ad presentan una enorme desventaja ya que, por su tamaño, despiertan una enorme respuesta inmune, tanto innata como adaptativa, en el organismo que los recibe, lo cual trae como consecuencia la producción de citoquinas proinflamatorias, como el factor de necrosis tumoral alfa (TNF-alfa), la interleuquina 6 (IL-6) y el interferón tipo 1 (IFN-alfa y beta) (antivirales por excelencia), que inducen su eliminación.
Entrada de Ad a la célula
La vía de entrada del Ad a la célula huésped es por el mecanismo de endocitosis mediada por receptor, en el que las integrinas de la membrana celular desempeñan un papel muy importante. Los dos receptores que los Ad reconocen para introducirse a las células son el receptor CD46 para el serotipo B y el receptor específico de Ad (CAR) para los demás serotipos. El receptor CAR interactúa con la fracción globular knob del Ad, mientras que las integrinas (alfav-beta3 y alfav-beta5) interactúan con la base de la pentona.7 El Ad entra a la célula en un endosoma cuya acidificación favorece la disociación de la cápside, liberando el ADN viral al citoplasma, el cual es transportado con ayuda de los microtúbulos de dineína al núcleo, donde permanece de forma episomal, es decir, no se integra al ADN del huésped sino que se mantiene como un fragmento independiente (Fig.1).8,9
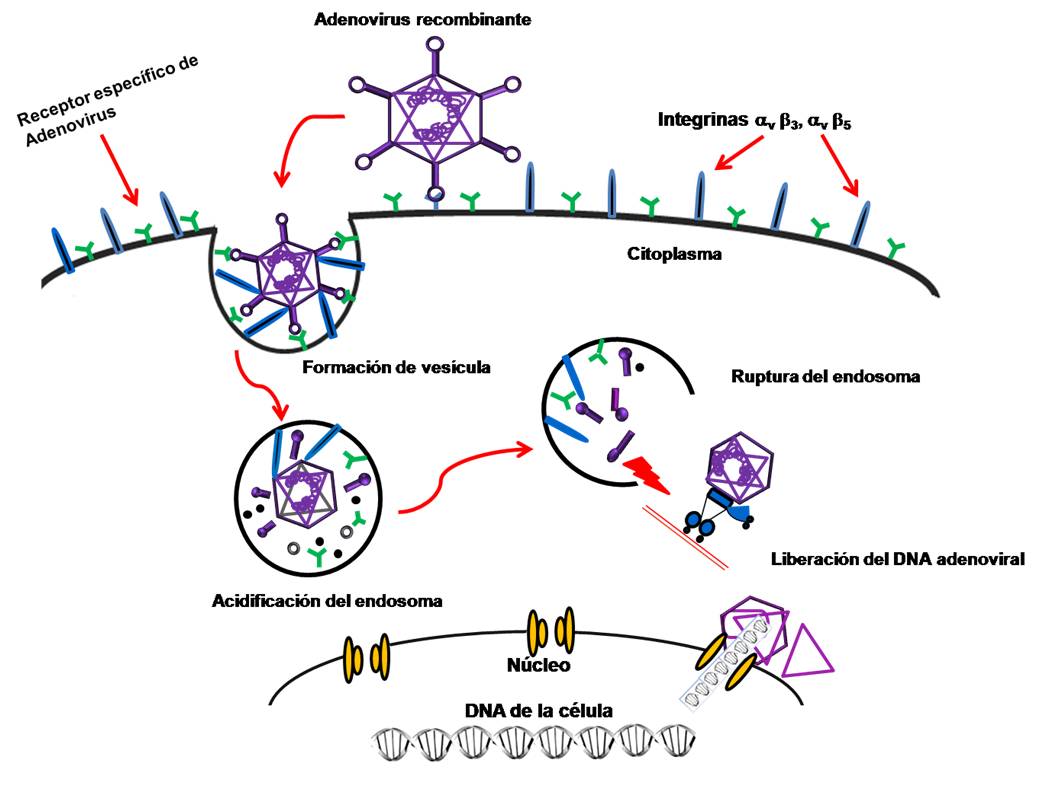
Respuesta inmune innata hacia los Ad
Los Ad inducen una fuerte respuesta inmunológica en la que la inmunidad innata, orquestada por macrófagos y células dendríticas, es el principal mecanismo para su eliminación. La administración sistémica de Ad permite la asociación de éstos con los factores de la coagulación VIII, IX y X, así como con la proteína C-reactiva. Esta asociación promueve la transducción de hepatocitos y de células de Kupffer, los cuales, mediante las vías de señalización de la proteína quinasa activada por mitógeno (MAPK) y del factor nuclear kappa beta (NFk-b), responden secretando citoquinas proinflamatorias como IL-6, IL-8, IL-12, TNF-alfa e IFN-gamma, con lo cual se establece un proceso inflamatorio manifestado como daño hepático agudo que culmina con la eliminación de los Ad en las primeras 24 horas.10-14 Las moléculas más inmunogénicas son las proteínas de la cápside, el ADN y los transcriptos virales. Por su parte, los receptores tipo Toll reconocen al Ad de manera intracelular e inducen la expresión de IFN-alfa e IFN-beta,15 lo que conduce a una respuesta humoral y celular contra el vector y contra el transgen. Debido a esto, el IFN-alfa y el IFN-beta?han sido considerados como moléculas importantes en la eliminación de Ad.
Interferones tipo 1 en la inmunidad contra Ad
Los interferones son citoquinas clave en la respuesta inmune contra virus, bacterias, parásitos o células tumorales. Participan tanto en la inmunidad innata como en la adaptativa, y logran establecer un estado antiviral intracelular que evita la replicación del virus y su propagación a las células vecinas.
Existen 3 tipos de IFN: tipo 1, tipo 2 y tipo 3, de los cuales sólo el tipo 1 –representado por IFN-alfa e IFN-beta– tiene propiedades antivirales. La expresión de IFN tipo 1 induce la actividad transcripcional de IRF3 (interferon regulatory factor), el cual estimula la expresión de los genes antivirales proteína quinasa R, mixiovirus y el mismo IFN. Los niveles elevados de IFN estimulan tanto la respuesta inmune innata como la adaptativa contra Ad. Zhu y colaboradores demostraron que la administración de Ad en ratones deficientes del receptor para IFN tipo 1 no induce expresión de IL-6 ni de IL-12, por lo que no hay activación de células T; de esta forma queda en evidencia la importancia del IFN tipo 1 en la eliminación de Ad.15
Respuesta inmune adaptativa hacia Ad
La respuesta inmune adaptativa se produce semanas después de contacto del organismo con los Ad, la cual se caracteriza por una producción elevada de anticuerpos contra ellos, así como la inducción de linfocitos T citotóxicos CD8+ (LTC CD8+) contra proteínas del Ad o productos del transgen. Esta respuesta conduce al organismo a daño multiorgánico, dañando preferentemente el hígado.16-18 Después de la administración intravenosa de Ad, éstos son rápidamente captados por las células presentadoras de antígeno (APC, antigen-presenting cells), las cuales, por medio del complejo mayor de histocompatibilidad clase I (MHC I), presentan los antígenos virales a los linfocitos CD8+ induciendo una respuesta inmune celular. Los linfocitos CD8+ son los responsables de eliminar a las células transducidas, provocando también la pérdida de la expresión del gen terapéutico. La presentación de antígenos virales mediante el complejo mayor de histocompatibilidad clase II (MHC II) activa a las células CD4+ y a los linfocitos B, lo cual induce una respuesta inmune humoral con producción de anticuerpos anti-adenovirus. La presencia de estos anticuerpos neutraliza a las partículas virales circulantes, bloqueando la entrada a la célula y eliminándolas (Fig. 2).14,19,20
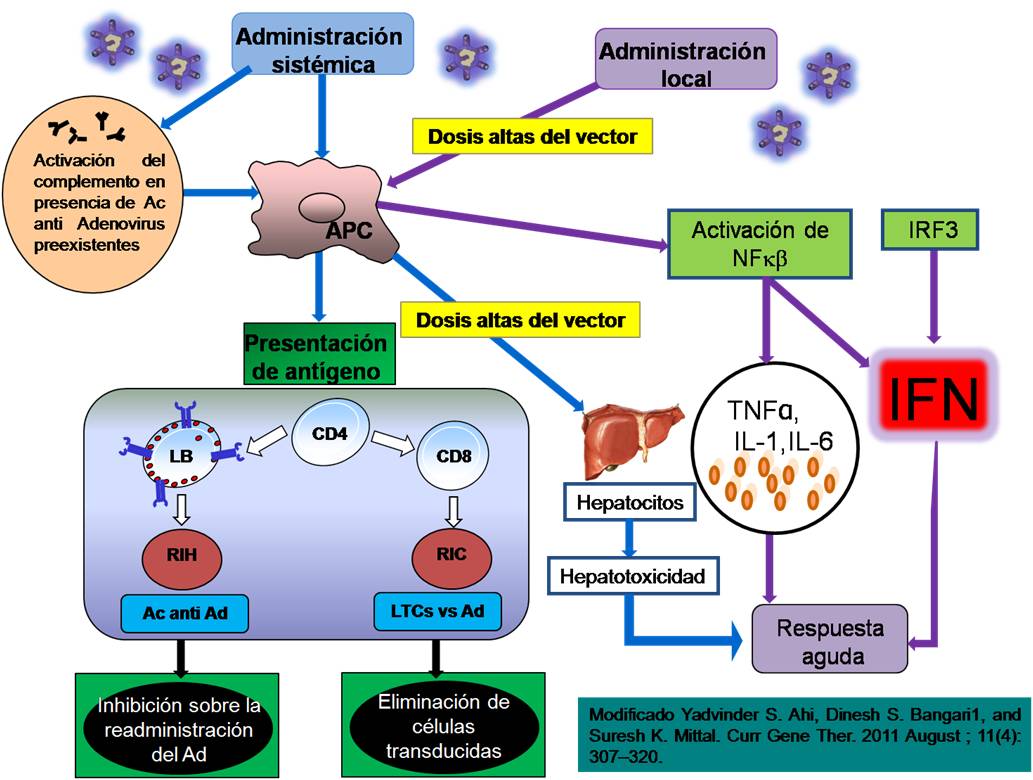
Estrategias utilizadas para evitar la inmunidad
Puesto que el principal inconveniente del uso de Ad en protocolos clínicos es la respuesta inmune que despiertan en el huésped, los investigadores se han dado a la tarea de diseñar estrategias que evadan esa respuesta inmune, tratando así de hacer más eficiente la tansducción a las células blanco y expresando por más tiempo el gen terapéutico. Algunas de estas estrategias han logrado resultados muy prometedores y son las que se presentan a continuación.
Krasnykh y cols. realizaron por primera vez la seudotipificación de la fibra de la cápside de los Ad, reemplazando el gen completo de la fibra o el dominio knob con el gen correspondiente de otro serotipo de Ad que infecta al ser humano y que reconoce un receptor celular diferente a CAR. Esta estrategia proporciona, además, un método rápido y sencillo de producción de rAd con fibras modificadas, que puede manipular el tropismo de los Ad a unas células en particular.21 Para ello, incorporaron ligandos en la fibra del Ad la cual medía la unión del Ad a su receptor en la célula. Utilizando proteínas recombinantes de la fibra expresadas en células de insecto infectadas con baculovirus, demostraron que la incorporación del octapéptido FLAG en el bucle de HI no realiza ablación de la trimerización de la fibra, no altera la unión del Ad a su receptor y no modifica sus funciones biológicas. A continuación se generó un Ad recombinante que contenía esta fibra modificada y se demostró que la secuencia del péptido corto era compatible con las funciones biológicas de la fibra. Esta información sugiriere que los ligandos heterólogos se pueden incorporar en el bucle HI de la perilla de la fibra y que esta localización tiene propiedades compatibles para su empleo como estrategia de reorientación de los Ad.22
Por su parte, Belousova y cols. identificaron el bucle HI del dominio perilla de la fibra como el sitio ideal para la incorporación de ligandos y sugirieron que las propiedades estructurales de este bucle permiten la inserción de una amplia variedad de ligandos, incluidos polipéptidos grandes. Mediante una serie de estudios se demostró que la fibra HI del Ad5 tolera inserciones de péptidos de hasta 100 aminoácidos, con efectos mínimos sobre la integridad del virión, lo que sugiere que es posible insertar un péptido en un ligando sin alterar su plegamiento ni su unión al receptor.23 Esta estrategia permite realizar una segunda administración con un rAd modificado en su fibra que conserva la misma afinidad por el receptor, sin correr el riesgo de que los anticuerpos producidos contra el primer Ad lo eliminen.
Havenga y cols. construyeron rAd tipo 5 capaces de infectar células humanas reconociendo receptores diferentes a CAR. Investigaron la diversidad natural del tropismo tisular de los Ad humanos y descubrieron que llevan fibras procedentes de otros serotipos y pueden infectar células humanas de estirpes diferentes a las convencionales (células dendríticas, endoteliales, de músculo liso, fibroblastos, de médula ósea), lo que permite proponer nuevos enfoques de tratamiento en terapia génica para diversas enfermedades.24
En otra estrategia para disminuir la inmunogenicidad de los Ad, Dimitriev y cols. incorporaron una secuencia de residuos Arg-Gly-Asp en el bucle HI de la fibra knob para utilizar un receptor alternativo durante el proceso de transducción, con la finalidad de poder transducir eficazmente células tumorales primarias. Con esta estrategia, estos investigadores lograron incrementar la transferencia de genes en una línea celular de cáncer de ovario mediante un mecanismo independiente de CAR.25
Koizumi y cols. informaron que la transducción en ratones C57BL6 con Ad modificados genéticamente en la fibra knob (al que se le incorporó un péptido de polilisina) indujo niveles más bajos de IL-6 y alanino aminotransferasa en suero que el vector convencional después de una administración intravenosa, lo que permitió al mismo tiempo un incremento en la expresión del transgen.26
El uso de moléculas de ARN de interferencia (siARN) como estrategia anti-adenoviral también ha sido probado. En un estudio realizado por Qingfeng y cols. se administró un siARN contra el ARNm de la quimioquina fractalquina en hígados de ratas antes de ser transducidas con Ad. Esto evitó el daño hepático agudo originado por la administración del vector adenoviral. El infiltrado inflamatorio de células natural killer fue significativamente menor, así como los niveles séricos de IFN-gamma, con una prolongación en el tiempo de expresión del transgen.12
Zhu y cols. demostraron que la administración de anticuerpos anti-IFN-alfa e IFN-beta en ratones C57BL/6 6 horas antes de la transducción de Ad-LacZ disminuyó drásticamente la respuesta inmune, tanto innata como adaptativa, hacia los vectores adenovirales. Los ratones tratados con anticuerpos anti-IFN-alfa e IFN-beta tuvieron un aumento en el número de copias de ADN adenoviral, mayor estabilidad del transgen y reducción en la activación de células T y en la inflamación hepática, tres días después de la administración. Estos resultados permitieron a los autores sugerir que el bloqueo de IFN tipo 1 podría ser útil en protocolos de terapia génica cuando se utilicen Ad como vectores, lo que permite la expresión del gen terapéutico durante más tiempo.15
El uso de polietilenglicol catiónico, de ácido láctico y de ácido glicólico para cubrir a los Ad mediante encapsulación con polímeros, también ha mostrado ser una estrategia eficaz para evadir a los anticuerpos neutralizantes contra los vectores adenovirales.27,28 La microencapsulación de AdH5 recombinante eludió de manera eficaz la respuesta inmune en ratones inmunizados, vía intranasal o intraperitoneal, con AdCA36lacZ, encapsulado dentro de micropartículas del polímero alginato.29
Por otra parte, también se han utilizado Ad encapsulados con microesferas de polientilenglicol expresando GFP, con la finalidad de probar la eficacia de transducción con esta estrategia. Los Ad encapsulados incrementaron en un 23% la transducción respecto a los no encapsulados, lo que demostró que el uso de PLG como agente encapsulante es una alternativa eficaz para el empleo de Ad en terapia génica.30
Otra estrategia para evadir la respuesta inmune es la utilización de un serotipo de Ad diferente al 2 y al 5, como es el subgrupo B, el cual reconoce un cofactor de unión a la membrana diferente a CAR denominado CD46. La unión a CD46 no desencadena la respuesta inmune celular que se presenta cuando la unión es a través de CAR, por lo que su empleo resulta prometedor cuando se pretende evadir la inmunidad.31 Es importante mencionar que al ser los Ad vectores que no se integran al genoma de la célula sino que quedan de manera episomal en el núcleo, no representan riesgo alguno de mutación insercional por lo que su uso no está asociado con la aparición de tumores o cáncer.8 Los vectores Ad no humanos también son utilizados para evadir la inmunidad preexistente, lo que permite que más Ad ingresen a sus células blanco.32
La construcción de vectores Ad Helper-Depent y su uso en terapia génica ha eliminado el problema de la expresión génica viral residual asociada con los vectores de primera generación.33 Estudios iniciales en animales de laboratorio han demostrado que este tipo de Ad provoca una respuesta inmune limitada en las células, permitiendo la expresión del transgen durante más tiempo.34 Estos vectores tiene una gran variedad de aplicaciones; sin embargo, su producción aún es más compleja que la de los Ad de primera generación.35 Su gran capacidad de inserción y la baja respuesta celular hace de estos vectores una estrategia prometedora para terapia génica de enfermedades que requieran la expresión de un gen terapéutico de manera temporal.36
Finalmente, es importante mencionar el uso de Ad en protocolos de terapia génica dirigida contra el cáncer. En este tipo de enfermedades, la respuesta inmune inducida por los Ad, y en especial hacia las células transducidas, permite la eliminación de células cancerosas por lo que es utilizada como estrategia de tratamiento para diferentes tipos de cánceres. En protocolos experimentales y clínicos de cáncer de próstata y de ovario, el envío de Ad que contienen el gen suicida timidina quinasa en combinación con quimioterapia incrementó de manera significativa el número de células cancerosas que mueren por apoptosis o necrosis.37-39
Conclusión
Los adenovirus son, en la actualidad, los vectores más utilizados en ensayos clínicos de terapia génica. A pesar de que su administración induce una respuesta inmunológica elevada, se prefieren sobre otros vectores pues presentan enormes ventajas, como su producción relativamente sencilla y su manejo que no representa ningún riesgo para la salud. Los estudios realizados para tratar de entender los mecanismos moleculares por los cuales los Ad transducen a las células y la manera en que despiertan una respuesta inmune en el huésped son de gran importancia. Los conocimientos actuales sobre estos mecanismos han permitido proponer estrategias de evasión inmunológica con la finalidad de lograr un efecto terapéutico más prolongado. La superación de los obstáculos que representa la respuesta inmune y la dirección del vector hacia un órgano específico son los grandes retos a vencer. Cuando logremos que los Ad alcancen su célula blanco, expresen su gen terapéutico y el organismo permita nuevas administraciones del vector, se habrá realizado un gran avance en el tratamiento de enfermedades, no sólo genéticas sino también crónico-degenerativas, con lo cual se logrará incrementar las expectativas de vida de muchos pacientes que esperan el desarrollo de nuevas estrategias terapéuticas a los males que los aquejan.
Bibliografía del artículo
1. Assessment of adenoviral vector safety and toxicity: report of the National Institutes of Health Recombinant DNA Advisory Committee. Hum Gene Ther 13(1):3-13, 2002.
2. Coughlan L, Alba R, Parker AL, Bradshaw AC, McNeish IA, Nicklin SA, et al. Tropism-modification strategies for targeted gene delivery using adenoviral vectors. Viruses 2(10):2290-355, 2010.
3. Wiley J. The journal of Gene Medicine; 2014.
4. Arnberg N. Adenovirus receptors: implications for tropism, treatment and targeting. Rev Med Virol 19(3):165-q78, 2009.
5. Mowa MB, Crowther C, Arbuthnot P. Therapeutic potential of adenoviral vectors for delivery of expressed RNAi activators. Expert Opin Drug Deliv 7(12):1373-1385, 2010.
6. Kay MA, Glorioso JC, Naldini L. Viral vectors for gene therapy: the art of turning infectious agents into vehicles of therapeutics. Nat Med 7(1):33-40, 2001.
7. Glasgow JN, Everts M, Curiel DT. Transductional targeting of adenovirus vectors for gene therapy. Cancer Gene Ther 13(9):830-844, 2006.
8. Trotman LC, Mosberger N, Fornerod M, Stidwill RP, Greber UF. Import of adenovirus DNA involves the nuclear pore complex receptor CAN/Nup214 and histone H1. Nat Cell Biol 3(12):1092-1100, 2001.
9. Suomalainen M, Nakano MY, Keller S, Boucke K, Stidwill RP, Greber UF. Microtubule-dependent plus- and minus end-directed motilities are competing processes for nuclear targeting of adenovirus. J Cell Biol 144(4):657-672, 1999.
10. Schnell MA, Zhang Y, Tazelaar J, Gao GP, Yu QC, Qian R, et al. Activation of innate immunity in nonhuman primates following intraportal administration of adenoviral vectors. Mol Ther 3(5):708-722, 2001.
11. Taniguchi M, Seino K, Nakayama T. The NKT cell system: bridging innate and acquired immunity. Nat Immunol 4(12):1164-1165, 2003.
12. Chen Q, Wei H, Sun R, Zhang J, Tian Z. Therapeutic RNA silencing of Cys-X3-Cys chemokine ligand 1 gene prevents mice from adenovirus vector-induced acute liver injury. Hepatology 47(2):648-658, 2008.
13. Guidotti LG, Chisari FV. Noncytolytic control of viral infections by the innate and adaptive immune response. Annu Rev Immunol 19:65-91, 2001.
14. Ahi YS, Bangari DS, Mittal SK. Adenoviral vector immunity: its implications and circumvention strategies. Curr Gene Ther 11(4):307-320, 2011.
15. Zhu J, Huang X, Yang Y. Innate immune response to adenoviral vectors is mediated by both Toll-like receptor-dependent and -independent pathways. J Virol 81(7):3170-3180, 2007.
16. Liu Q, Muruve DA. Molecular basis of the inflammatory response to adenovirus vectors. Gene Ther 10(11):935-940, 2003.
17. Jooss K, Chirmule N. Immunity to adenovirus and adeno-associated viral vectors: implications for gene therapy. Gene Ther 10(11):955-963, 2003.
18. Chen D, Murphy B, Sung R, Bromberg JS. Adaptive and innate immune responses to gene transfer vectors: role of cytokines and chemokines in vector function. Gene Ther 10(11):991-998, 2003.
19. Bangari DS, Mittal SK. Current strategies and future directions for eluding adenoviral vector immunity. Curr Gene Ther 6(2):215-226, 2006.
20. Thaci B, Ulasov IV, Wainwright DA, Lesniak MS. The challenge for gene therapy: innate immune response to adenoviruses. Oncotarget 2(3):113-121, 2011.
21. Krasnykh VN, Mikheeva GV, Douglas JT, Curiel DT. Generation of recombinant adenovirus vectors with modified fibers for altering viral tropism. J Virol 70(10):6839-6846, 1996.
22. Krasnykh V, Dmitriev I, Mikheeva G, Miller CR, Belousova N, Curiel DT. Characterization of an adenovirus vector containing a heterologous peptide epitope in the HI loop of the fiber knob. J Virol 72(3):1844-1852, 1998.
23. Belousova N, Krendelchtchikova V, Curiel DT, Krasnykh V. Modulation of adenovirus vector tropism via incorporation of polypeptide ligands into the fiber protein. J Virol 76(17):8621-8631, 2002.
24. Havenga MJ, Lemckert AA, Ophorst OJ, van Meijer M, Germeraad WT, Grimbergen J, et al. Exploiting the natural diversity in adenovirus tropism for therapy and prevention of disease. J Virol 76(9):4612-4620, 2002.
25. Dmitriev I, Krasnykh V, Miller CR, Wang M, Kashentseva E, Mikheeva G, et al. An adenovirus vector with genetically modified fibers demonstrates expanded tropism via utilization of a coxsackievirus and adenovirus receptor-independent cell entry mechanism. J Virol 72(12):9706-9713, 1998.
26. Koizumi N, Yamaguchi T, Kawabata K, Sakurai F, Sasaki T, Watanabe Y, et al. Fiber-modified adenovirus vectors decrease liver toxicity through reduced IL-6 production. J Immunol 178(3):1767-1773, 2007.
27. Chillon M, Lee JH, Fasbender A, Welsh MJ. Adenovirus complexed with polyethylene glycol and cationic lipid is shielded from neutralizing antibodies in vitro. Gene Ther 5(7):995-1002, 1998.
28. Beer SJ, Matthews CB, Stein CS, Ross BD, Hilfinger JM, Davidson BL. Poly (lactic-glycolic) acid copolymer encapsulation of recombinant adenovirus reduces immunogenicity in vivo. Gene Ther 5(6):740-746, 1998.
29. Sailaja G, HogenEsch H, North A, Hays J, Mittal SK. Encapsulation of recombinant adenovirus into alginate microspheres circumvents vector-specific immune response. Gene Ther 9(24):1722-1729, 2002.
30. Turner P, Petch A, Al-Rubeai M. Encapsulation of viral vectors for gene therapy applications. Biotechnol Prog 23(2):423-429, 2007.
31. Gaggar A, Shayakhmetov DM, Lieber A. CD46 is a cellular receptor for group B adenoviruses. Nat Med 9(11):1408-1412, 2003.
32. Bangari DS, Mittal SK. Development of nonhuman adenoviruses as vaccine vectors. Vaccine 24(7):849-862, 2006.
33. Parks RJ, Chen L, Anton M, Sankar U, Rudnicki MA, Graham FL. A helper-dependent adenovirus vector system: removal of helper virus by Cre-mediated excision of the viral packaging signal. Proc Natl Acad Sci USA 93(24):13565-13570, 1996.
34. Morsy MA, Gu M, Motzel S, Zhao J, Lin J, Su Q, et al. An adenoviral vector deleted for all viral coding sequences results in enhanced safety and extended expression of a leptin transgene. Proc Natl Acad Sci USA 95(14):7866-7871, 1998.
35. Kreppel F. Production of high-capacity adenovirus vectors. Methods Mol Biol 1089:211-229, 2014.
36. Cots D, Bosch A, Chillon M. Helper dependent adenovirus vectors: progress and future prospects. Curr Gene Ther 13(5):370-381, 2013.
37. Ebara S, Shimura S, Nasu Y, Kaku H, Kumon H, Yang G, et al. Gene therapy for prostate cancer: toxicological profile of four HSV-tk transducing adenoviral vectors regulated by different promoters. Prostate Cancer Prostatic 5(4):316-325, 2002.
38. Kieback DG, Fischer DC, Engehausen DG, Sauerbrei W, Oehler MK, Tong XW, et al. Intraperitoneal adenovirus-mediated suicide gene therapy in combination with either topotecan or paclitaxel in nude mice with human ovarian cancer. Cancer Gene Ther 9(5):478-481, 2002.
39. Rojas-Martinez A, Manzanera AG, Sukin SW, Esteban-Maria J, Gonzalez-Guerrero JF, Gomez-Guerra L, et al. Intraprostatic distribution and long-term follow-up after AdV-tk immunotherapy as neoadjuvant to surgery in patients with prostate cancer. Cancer Gene Ther 20(11):642-649, 2013.
|
|
©
Está
expresamente prohibida la redistribución y la redifusión de todo o parte de los
contenidos de la Sociedad Iberoamericana de Información Científica (SIIC) S.A. sin
previo y expreso consentimiento de SIIC |

|
|

